





1.染色体の構築
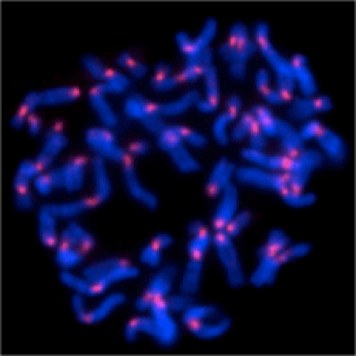
細胞が分裂する際には、クロマチン線維は凝縮して「染色体」に変換され、その染色体を分配することで、細胞は過不足なくゲノムを受け継いでいます。私たちは、染色体が凝縮する過程に、どのような分子が関与しているか?それら分子の動態はどのように制御されているか?ということに興味を持っています。特に、正確な染色体の分離に必須であることが知られている、コンデンシンおよびコヒーシンの機能はこの二つの因子にクローズアップしその制御メカニズムを追究しています。
2.染色体と微小管の結合制御
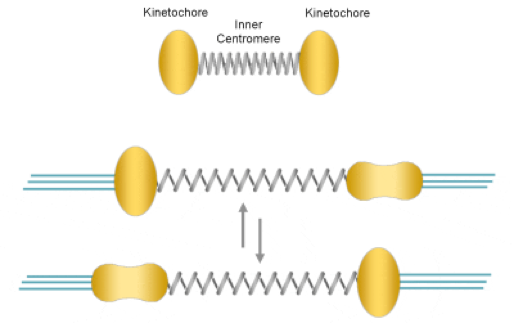
正確な染色体の分配を達成するためには、染色体と微小管の結合制御(姉妹染色分体がそれぞれ一方の紡錘体極と結合すること)と、染色体分離のタイミング制御(全ての染色体において正しい結合が完了した後に姉妹染色分体をいっせいに分離すること)が鍵であり、それぞれ、微小管の誤接続を解除する “修正機構” と、後期の開始を抑制する “紡錘体形成チェックポイント” という機能に依存していることが知られています。私たちの研究室では、主として生細胞の観察技術によって、これらの細胞機能を深く調べています。
3.染色体分離のタイミング制御
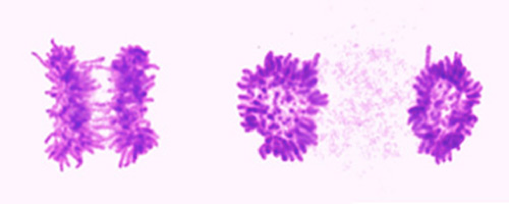
正確な染色体分離は、正確な動原体と微小管の結合によって達成され、結合の成否は紡錘体チェックポイント機構によっても厳重に監視されている。しかし、紡錘体チェックポイントの検問点を通過しても、完全に正しい分配をするためには、染色体は「いっせいに」分離しなければならない可能性があると考えています。私たちは「染色体分離の同期性」に着目し、どのような仕掛けによってこの同期性が保証されているのか、そのメカニズムを探究しています。
4.がん細胞における染色体不安定性の発生機構
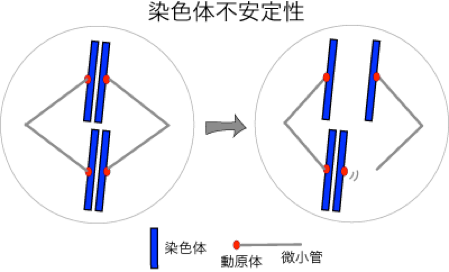
がんが悪性である最大の理由は、あらゆる治療戦略でがん細胞の除去を試みても、腫瘍は多様な性質を獲得した細胞から構成されるために、がん細胞を残さずに根絶することが難しいからです。昔から多くのがんはそれを構成するそれぞれの細胞の染色体数が異なる異数性であることが知られ、がん細胞は、細胞分裂の度に染色体数が変動する染色体不安定性と呼ばれる病態に陥っていると考えられます。実際に、異数性細胞の出現と臨床的な悪性度とは相関がみられることからも、染色体不安定性の獲得とがん細胞の不均一性とは無関係ではないはずで、染色体不安定性の究明こそが、がん病態の根幹的な理解につながると考えます。